2026年6月,我院岳田利教授与西班牙维戈大学肖建波教授联合研究团队在国际期刊《Redox Biology》(Q1, IF=16.2)发表题为“Early-life redox perturbation programs intergenerational redox vulnerability through DAF-16/FOXO and SKN-1/Nrf2 signaling in Caenorhabditis elegans”的研究论文。第一作者为2022级博士生梁静一美。
蛋氨酸是一种人体必需氨基酸,也是连接一碳代谢、甲基供体生成、氧化还原稳态与表观遗传调控的重要代谢枢纽,常被认为是维持生长发育和细胞功能所必需的营养物质。
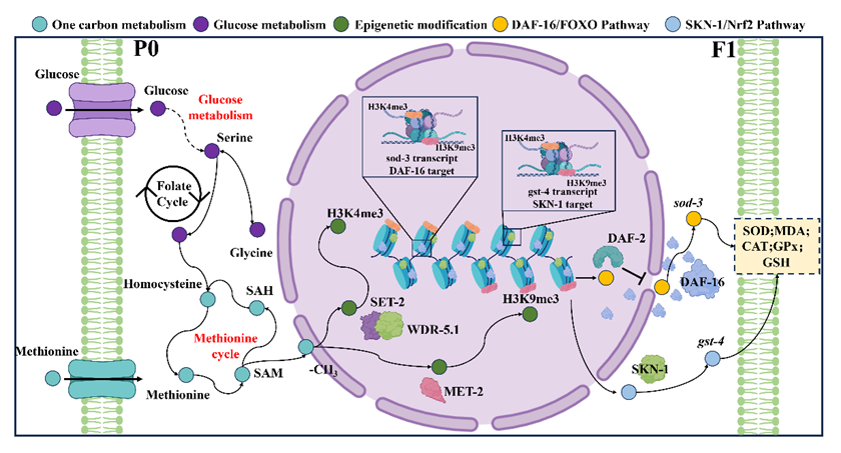
研究发现,亲代早期蛋氨酸过量可通过一碳代谢与表观遗传调控的耦联,将代谢压力传递至未直接暴露的子代,削弱其氧化还原韧性并加速功能衰退。通过建立亲代不同生殖阶段的蛋氨酸暴露模型,研究团队发现,早期生殖阶段是最敏感的时间窗口。该阶段的蛋氨酸过量可导致F1代子代运动能力下降、寿命缩短、氧化应激升高和抗氧化防御能力减弱,且损伤程度随蛋氨酸剂量增加而加重。停止暴露后,相关异常在F2代中基本恢复,说明单次暴露主要引起直接的代际编程效应;而连续多代暴露则会使运动、寿命和氧化还原损伤逐代累积。为追踪这一代际损伤的代谢来源,研究团队结合UHPLC-HRMS非靶向代谢组学、GC-MS和LC-MS靶向定量以及U-¹³C₆-葡萄糖稳定同位素示踪进行分析。结果显示,亲代蛋氨酸过量促使子代葡萄糖来源碳流向丝氨酸-甘氨酸一碳代谢通路重新分配,并扰乱糖酵解和三羧酸循环。与此同时,子代体内蛋氨酸、S-腺苷同型半胱氨酸和同型半胱氨酸积累,而S-腺苷甲硫氨酸及SAM/SAH比值下降,说明过量蛋氨酸并未增加可利用的甲基供体,反而造成蛋氨酸循环超负荷和甲基化潜能受损。
这种甲基供体失衡进一步引起子代染色质重塑,表现为H3K4me3升高和H3K9me3降低,同时伴随SET-2/WDR-5.1异常激活和MET-2抑制。通过set-2、wdr-5.1和met-2缺陷型线虫验证,研究团队证明SET-2/WDR-5.1依赖的H3K4过度甲基化是推动子代功能衰退的重要环节。转录组和遗传学实验进一步表明,染色质异常最终限制了DAF-16/FOXO和SKN-1/Nrf2应激防御通路,抑制sod-3和gst-4等抗氧化基因表达,使子代难以充分启动自身的应激适应程序。过氧化氢对照实验显示,单纯氧化应激不能复制上述组蛋白甲基化异常和子代持续损伤,说明真正的上游驱动力是一碳代谢失衡、甲基供体紊乱及其引起的染色质重塑。
研究团队还发现,茶多酚表没食子儿茶素没食子酸酯(EGCG)能够有效阻断这一过程。EGCG可剂量依赖性地改善子代运动能力、延长寿命、增强抗氧化防御并降低脂质过氧化,其中与蛋氨酸同时干预的效果优于提前干预。进一步分析表明,EGCG能够纠正葡萄糖碳流向一碳代谢通路的异常偏移,降低蛋氨酸、SAH和同型半胱氨酸积累,恢复SAM及SAM/SAH稳态;同时抑制SET-2/WDR-5.1异常激活,恢复MET-2表达,纠正H3K4me3/H3K9me3失衡,并重新激活DAF-16/FOXO和SKN-1/Nrf2介导的抗氧化程序。
这项研究表明,EGCG的保护作用并非仅源于直接清除自由基,而是通过协同调节蛋氨酸代谢、甲基供体稳态、组蛋白甲基化和应激转录网络,从源头阻止亲代代谢压力转化为子代持续的氧化还原脆弱性。研究为理解生命早期营养环境如何塑造子代长期健康提供了新的理论依据,也为利用膳食多酚开展早期精准营养干预提供了新的研究方向。

原文链接:https://doi.org/10.1016/j.redox.2026.104168

 关注我们
关注我们
 友情链接
友情链接 学校首页
学校首页 快捷导航
快捷导航